遺伝子連鎖(遺伝的連鎖)とは?定義・クロスオーバーと遺伝地図の仕組み
遺伝子連鎖の定義からクロスオーバー、遺伝地図の作成法まで図解でわかりやすく解説。距離計算やm.u.の意味も丁寧に紹介。
遺伝的連鎖は、異なる遺伝子座の対立遺伝子がランダムに偏析しないときに起こります。このようにしてメンデルの第二法則は破られる。
遺伝子は同じ染色体上にあるときにつながっています。遺伝子は同じ染色体上にあるときに連結され、減数分裂の間も一緒にいる傾向があります。異なる染色体上にある遺伝子の対立遺伝子は、減数分裂中に染色体が独立して配列されるため、通常は連結されない。
減数分裂の際には、染色体が分離する際に DNA の交叉があります。そのため、同じ染色体上の対立遺伝子が分離して、異なる娘細胞に行くことがあります。対立遺伝子が染色体上で離れている場合は、対立遺伝子間でクロスオーバーが起こる可能性が高くなるため、このようなことが起こる可能性が高くなります。2つの遺伝子間の相対的な距離は、2つのリンクした遺伝的形質を示す生物の子孫を使って計算することができます。2 つの形質が一緒に走らない子孫の割合が記されています。両方の形質を示す子孫の割合が高いほど、2つの遺伝子が染色体上で近いことになります。
これは、染色体上の遺伝子をマッピングするために使用された最初の技術です。組換え体の数を計算することで、遺伝子間の距離の尺度を得ることができます。この距離は、遺伝地図単位(m.u.)またはセンチモルガンと呼ばれ、100分の1の減数分裂の産物が組換え体である遺伝子間の距離として定義されます。リンケージマップは、同じ染色体上に存在するいくつかの形質間のマップ距離を見つけることによって作成され、理想的には、複数のクロスオーバーの可能性を避けるために、形質間の大きなギャップを避けることができます。
画像ギャラリー
1 画像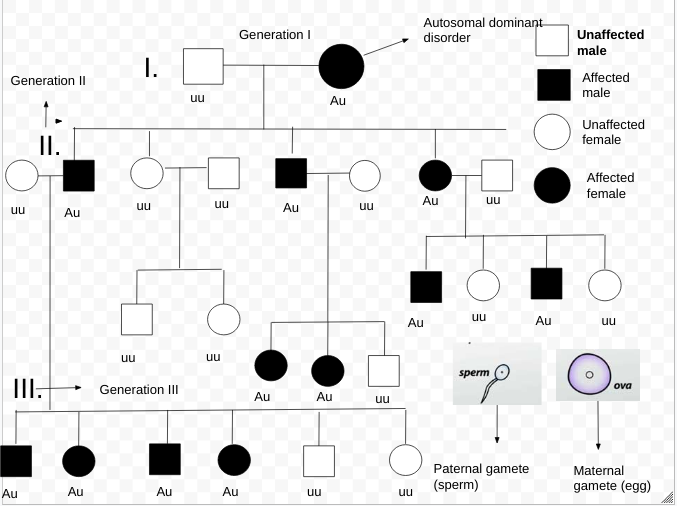
遺伝的連鎖の補足説明
遺伝的連鎖(linkage)とは、複数の遺伝子が同じ染色体上に位置するために、減数分裂での分離が独立に起こらず、特定の対立遺伝子の組合せが親の世代に近い形で子に伝わる現象です。連鎖が強いほど、親と同じ形質の組合せが子にもよく現れます。
クロスオーバー(組換え)の仕組み
- 減数分裂の前期に、相同染色体(父由来と母由来の対)は会合し、DNA鎖の一部が交換されます(交叉);これをクロスオーバーまたは組換えと呼びます。
- クロスオーバーが起きると、同一染色体上にある遺伝子の組合せが入れ替わり、新しい対立遺伝子の組合せをもつ配偶子が生じます。
- クロスオーバー確率は、一般に遺伝子間の物理的距離が大きいほど高くなります。したがって、遠く離れた遺伝子ほど分離しやすく、近接している遺伝子ほど連鎖が強く保たれます。
遺伝地図(リンケージマップ)の作成と計算
遺伝地図は、形質(またはマーカー)間の組換え頻度を基に作成されます。基本的な手順と考え方は次の通りです。
- 交配を行い、両親で異なる対立遺伝子の組合せがどの程度子に現れるか(再組換え体の割合)を数えます。
- 組換え頻度(recombination frequency, RF)=(再組換え個体数 / 総個体数)×100。これがパーセントで表され、概ねセンチモルガン(cM, 同義で m.u.)に対応します。例えば、RF = 10% は約10 cM。
- 注意:RF は実際の物理距離(塩基対の長さ)ではなく、「遺伝的距離」です。複数の交叉(多重交叉)が起きると、観察される組換え頻度は実際の距離を過小評価することがあります。
実務上の注意点:多重交叉とマッピング関数
- 2つのマーカー間に複数回のクロスオーバーが起こると、偶数回の交叉は再組換えを打ち消すため、観察される組換え率が実際の距離を過小評価します。
- そのため、実際の遺伝地図作成では Haldane や Kosambi のような「マッピング関数」を用いて、観察された組換え頻度から真の遺伝距離を推定します。これらの関数は多重交叉の確率を数学的に補正します。
- 遺伝地図上での距離が約50 cM を超えると、2つの遺伝子は実質的に独立に分配される(連鎖しているかどうか判別できない)ため、50 cM に近いまたはそれ以上の距離は「非連鎖」と見なされることがあります。
- 交叉の発生には「干渉(interference)」と呼ばれる効果があり、ある位置で交叉が起きると近傍で別の交叉が起こりにくくなる(正の干渉)ことがあります。干渉の存在は二重交叉の期待値を変え、マップ作成に影響します。
実験デザインと代表的手法
- テストクロス(testcross)やバッククロスは、再組換え頻度を測るための一般的な方法です。片方の親を既知のホモ接合にしておくことで、子に現れる対立遺伝子の組合せを明確に判定できます。
- 分子マーカー(SNP、SSR、RFLP など)の出現を用いれば、形質に限らず細かい遺伝地図を作成できます。現代では高密度SNPアレイや次世代シーケンシングにより、高解像度の地図が可能です。
- 遺伝地図と物理地図(ゲノム配列に基づく塩基対距離)は必ずしも一致しません。再組換えはゲノム上で不均一に発生するため、遺伝的距離と物理的距離の比は領域ごとに異なります。
応用と意義
- 遺伝子の位置同定(形質の連鎖解析、QTL解析)や品種改良におけるマーカー支援選抜(MAS)に用いられます。
- 染色体構造の研究や進化学的な研究にも重要で、種間での染色体再配列や交叉頻度の変化を通じて進化の痕跡をたどれます。
まとめ(ポイント)
- 遺伝的連鎖は同一染色体上の遺伝子が独立に分離しない現象で、メンデルの第二法則が当てはまらない例を説明する。
- クロスオーバーにより再組換えが起こり、その頻度から遺伝地図(cM)を作成できる。
- 多重交叉や干渉、物理距離との不一致など、遺伝地図には限界や補正が必要である。
質問と回答
Q:遺伝的連鎖とは何ですか?
A:遺伝的連鎖は、異なる遺伝子座の対立遺伝子がランダムに分離しない場合に起こり、メンデルの第二法則を破ります。遺伝子は同じ染色体上にあり、減数分裂の際に一緒になる傾向があるため、連鎖します。
Q:同じ染色体上の対立遺伝子は、どのように分離するのですか?
A:同じ染色体上の対立遺伝子は、減数分裂の際にDNAが交差することによって分離することがあります。染色体上の対立遺伝子が離れていると、交差が起こりやすいので、このような現象が起こる確率が高くなります。
Q:染色体上の遺伝子のマッピングにはどのような技術が使われたのですか?
A: 染色体上の遺伝子をマッピングする最初の方法は、2つの遺伝形質が関連する生物の子孫を用いて、2つの遺伝子間の相対距離を計算することでした。2つの遺伝形質が一緒になっていない子孫の割合を記録し、その割合が高いほど、染色体上の遺伝子の位置が近いことを示しています。
Q:遺伝地図単位(m.u.)またはセンチモルガンとは何ですか?
遺伝地図単位(m.u.)、またはセンチモルガンとは、減数分裂の産物が100個に1個組み合わされる遺伝子間の距離と定義されます。組換え頻度(RF)が1%の場合、1m.u.に相当する。
Q: 連鎖地図の作り方は?
A:連鎖地図は、同じ染色体上に存在するいくつかの形質間の地図上の距離を求めることによって作成することができます。理想的には、複数のクロスオーバーが同時に発生しないように、形質間に大きなギャップがないようにすることです。
Q:連鎖地図の作成において、形質間に大きなギャップがある場合はどうなるのでしょうか?
A:連鎖地図の作成において、形質間のギャップが大きいと、複数のクロスオーバーが同時に起こる可能性が高くなり、染色体上の遺伝子の位置や距離をマッピングしても、不正確な結果になる可能性があります。
著者
AlegsaOnline.com 遺伝子連鎖(遺伝的連鎖)とは?定義・クロスオーバーと遺伝地図の仕組み Leandro Alegsa
URL: https://ja.alegsaonline.com/art/37995