鳥脚類(オルニトポッド)とは?草食恐竜の特徴・進化と代表種まとめ
鳥脚類(オルニトポッド)の特徴・進化・代表種をわかりやすく解説。咀嚼装置の発達や生態、イグアノドンやハドロサウルス等の名種を図解で紹介。
Ornithopodsは、鳥に抱かれた恐竜のinfraorderです。彼らは、小さな、二足歩行の実行放牧者として始まり、彼らは白亜紀の世界で草食動物の最も成功したグループの一つになるまで、サイズと数で成長しました。
特徴
鳥脚類は草食性に特化した形質を数多く示します。骨格・頭骨・歯列の構造が高度に適応しており、効率的に植物を摂取・処理する能力を獲得しました。以下に主な特徴をまとめます。
- くちばしと頬:前方に発達したくちばしで植物を切り取り、頬(頬肉様構造)で食べ物を口内に留めて噛み砕きます。
- 複数列の歯・歯列バッテリー:特にアヒルビル(ハドロサウルス類)は多列の歯を密に並べた「歯列バッテリー」を持ち、すり潰すように咀嚼できました。
- 高度な咀嚼運動:彼らの顎と歯は単に噛むだけでなく、摺り合わせるような複雑な動きをするものが出現しました。これは、進化の利点として非常に重要でした。爬虫類的な基本構造に比べて、爬虫類がこれまでに開発した中で最も洗練された咀嚼装置の一つと考えられています。効率よく噛むことで植物を細かくし、より速く消化されることを可能にしました。
- 歩行様式の多様化:初期の鳥脚類は主に二足歩行でしたが、イグアノドン類やハドロサウルス類の一部では前肢が発達して四足歩行も行い、移動・採食に応じて二足・四足を使い分ける「兼用形」のものが現れました。
- 頭部装飾:ラメボサウルス亜科などでは骨質や骨板で作られたクレスト(冠状突起)を持ち、これを共鳴器として鳴き声を作ったとする説や、視覚的な個体識別・求愛などの機能が提案されています。
画像ギャラリー
10 画像








進化と分布
鳥脚類は古い地層から見つかっており、白亜紀の繁栄がよく知られていますが、その起源はジュラ紀・白亜紀にかけての拡散を経て、多様化しました。彼らは世界的に分布し、南極を含むすべての大陸から化石が知られています(南半球では化石がやや希少ですが存在します)。化石記録や足跡化石は群れで行動していた可能性や長距離移動、季節移動の存在を示唆します。
鳥脚類の主要な成功要因の一つは、前述の咀嚼装置の改善だけでなく、被食圧や環境の多様化に適応して体のサイズや生活形態を変化させたことです。最終的に彼らは白亜紀末の大量絶滅イベント(K–Pg境界)によって他の非アビア恐竜とともに壊滅的な打撃を受け、絶滅しました(ただし鳥類を除く)。
生態・行動の知見
- 群れと社会性:足跡や大量の骨の集合から、多くの鳥脚類は群れで生活し、幼獣の世話や共同移動を行っていた可能性があります。Maiasauraの巣化石は「やさしい母トカゲ(good mother lizard)」というニックネームを生み、親による育雛の証拠を与えました。
- 食性:葉、茎、種子、時には低木や木の芽などを食べていたと考えられます。消化補助のために胃石(胃に入れた小石)を使った種も知られます。
- 移動力:軽快な二足走行から、四足での安定した移動まで種によって異なり、高速走行に適した後肢を持つものも存在しました。
代表種(例)
グループの内部分類は研究の進展により変わりますが、代表的な系統や属は以下の通りです。
- ハイプシロフォドン類(例:Hypsilophodon)— 小型で敏捷、典型的な二足歩行の初期型。
- イグアノドン類(例:Iguanodon, Camptosaurus)— 中大型で、前肢が発達し四足歩行も可能になったもの。
- アヒルビルズ(ハドロサウルス科、例:Parasaurolophus, Corythosaurus, Edmontosaurus, Maiasaura)— 歯列バッテリーやクレストなど高度に特殊化し、白亜紀後期に繁栄した代表群。
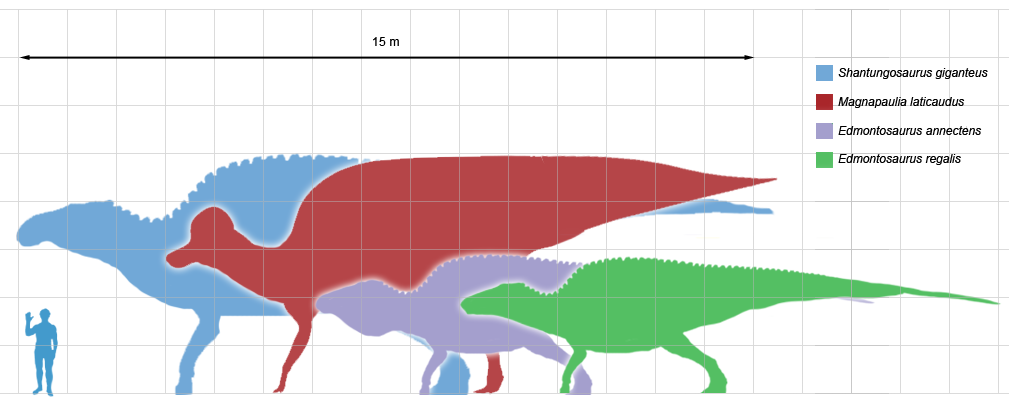
- その他(例:Dryosaurus, Tenontosaurus, Shantungosaurusなど)— 地域的に重要な大型・中型種を含む。
化石資料と研究
鳥脚類の化石は骨格だけでなく、足跡、巣跡、卵、そしてしばしば保存状態の良い頭骨や歯列が見つかるため、咀嚼機構や群れ行動、成長様式などの研究に有益です。特にハドロサウルス類の頭骨は内部構造がよく残り、共鳴器としての機能や嗅覚・聴覚の解剖学的解析が進んでいます。
まとめると、鳥脚類(オルニトポッド)は草食生活に極めて適応した恐竜群で、咀嚼装置の発達と歩行様式の多様化によりジュラ紀・白亜紀を通じて広く繁栄しました。角のある恐竜(セラトプシア)や頭の固い恐竜(パチケファロサウルス)の姉妹グループです。

説明
ライフスタイルと適応力
鳥脚類は白亜紀の世界で最も成功した草食動物のグループの一つとなり、景観を支配しました。彼らは、小さな、二足歩行のランニングブラウジングとして始まったが、サイズと数で成長した。イギリスのワイト島の白亜紀下部に生息するHypsilophodonはその典型的な例です。サイズが大きくなるにつれ、彼らは4本足で移動できるようになりましたが、食べ物にたどり着くため、あるいは身を守るため、あるいは走るために必要なときには、後ろ足で立ち上がる習慣が残っていました。
獣脚類の肉食動物に対する明らかな防御手段がほとんどなかったので、彼らの成功は驚くべきことかもしれません。彼らは走ることができ、群れの動物の通常の防御を持っていた。彼らは角張ったくちばしといくつかの軟骨の肋板を持っていた。Iguanadontsは、しかし、自分自身を守ることができました。彼らは非常に堅牢な(重くて強い)、そして短剣のような親指で武装していた。
初期の鳥脚類は約1メートル(3フィート)の長さが、おそらく非常に高速だった。吻足類のように硬い尻尾を持っていて、後ろ足で走るときにバランスを取るためのものでした。その後の鳥足類は、四つん這いで草を食べることに適応するようになり、その棘は湾曲し、バイソンのような現代の地上フィーダーの棘に似てきました。脊椎は湾曲しており、バイソンのような現代の地上飼育動物の脊椎に似ています。
その後の鳥脚類は大型化しましたが、首の長い、尾の長いサウロ脚類のような巨大な大きさにはなりませんでした。シャントゥンゴサウルスのような最大のものは、中型のサウナ脚類と同じくらいの重さがありましたが、15メートル(50フィート)を超えるような大きさにはなりませんでした。

分類
質問と回答
Q: 鳥脚類とは何ですか?
A:鳥脚類は、鳥類を飼う恐竜の下目です。
Q:鳥脚類はどのように進化したのですか?
A:鳥脚類は、二足歩行の小さな草食動物から始まり、白亜紀の世界で最も成功した草食動物のグループの1つになるまで、サイズと数を増やしていきました。
Q: 鳥脚類の主な進化的利点は何でしたか?
A:鳥脚類の進化的な利点は、爬虫類の中で最も洗練された咀嚼装置を発達させたことです。
Q: 鳥脚類の咀嚼器はどのような特徴を持っていたのですか?
A:くちばし、複数の歯列、頬袋、そして真の咀嚼を備えていました。
Q: 鳥脚類が効率よく咀嚼することの利点は何ですか?
A: 咀嚼が効率的であることは、物質をより小さく砕くことができるため、消化が早いということです。
Q: 鳥脚類の進化の頂点は何だったのでしょうか?
A:鳥脚類の進化の頂点は、アヒル口でした。
Q:鳥脚類の絶滅の原因は何ですか?
A:鳥脚類は白亜紀から第三紀にかけての絶滅イベントで、他の非鳥類恐竜と一緒に絶滅しました。
関連項目
著者
AlegsaOnline.com 鳥脚類(オルニトポッド)とは?草食恐竜の特徴・進化と代表種まとめ Leandro Alegsa
URL: https://ja.alegsaonline.com/art/73215
出典
- doi.org : 10.1016/j.cretres.2008.02.002
- doi.org : 10.1080/10292389209380426