渦鞭毛藻類とは|特徴・分類・生態 — 光合成・捕食・分布の解説
渦鞭毛藻類の特徴・分類・生態を図解でわかりやすく解説。光合成・捕食・共生・生物発光、最新推定約2,294種や海・淡水の分布も掲載。
渦鞭毛藻類は、鞭毛を持つ原生生物の大きなグループである。約半数は光合成を行い、残りはほとんどが他の原生生物を捕食する従属栄養生物である。光合成を行うものを「藻類」、捕食を行うものを「原生動物」と呼ぶことがある。
彼らは、異なる著者によって、目、クラス、門に分類されています。それらの関係については活発な研究が行われている。
ほとんどが海洋プランクトンですが、淡水にも生息しています。温度、塩分、水深に応じて分布しています。このグループには、多くの共生生物や生物発光種が含まれる。
海産の自由生活を営む渦鞭毛藻類は,現在約1,555種が記載されている。最新の推定では、海洋性、淡水性、寄生性の渦鞭毛藻を含めて、合計2,294種の現存する渦鞭毛藻があると考えられています。
画像ギャラリー
10 画像








特徴(形態と生理)
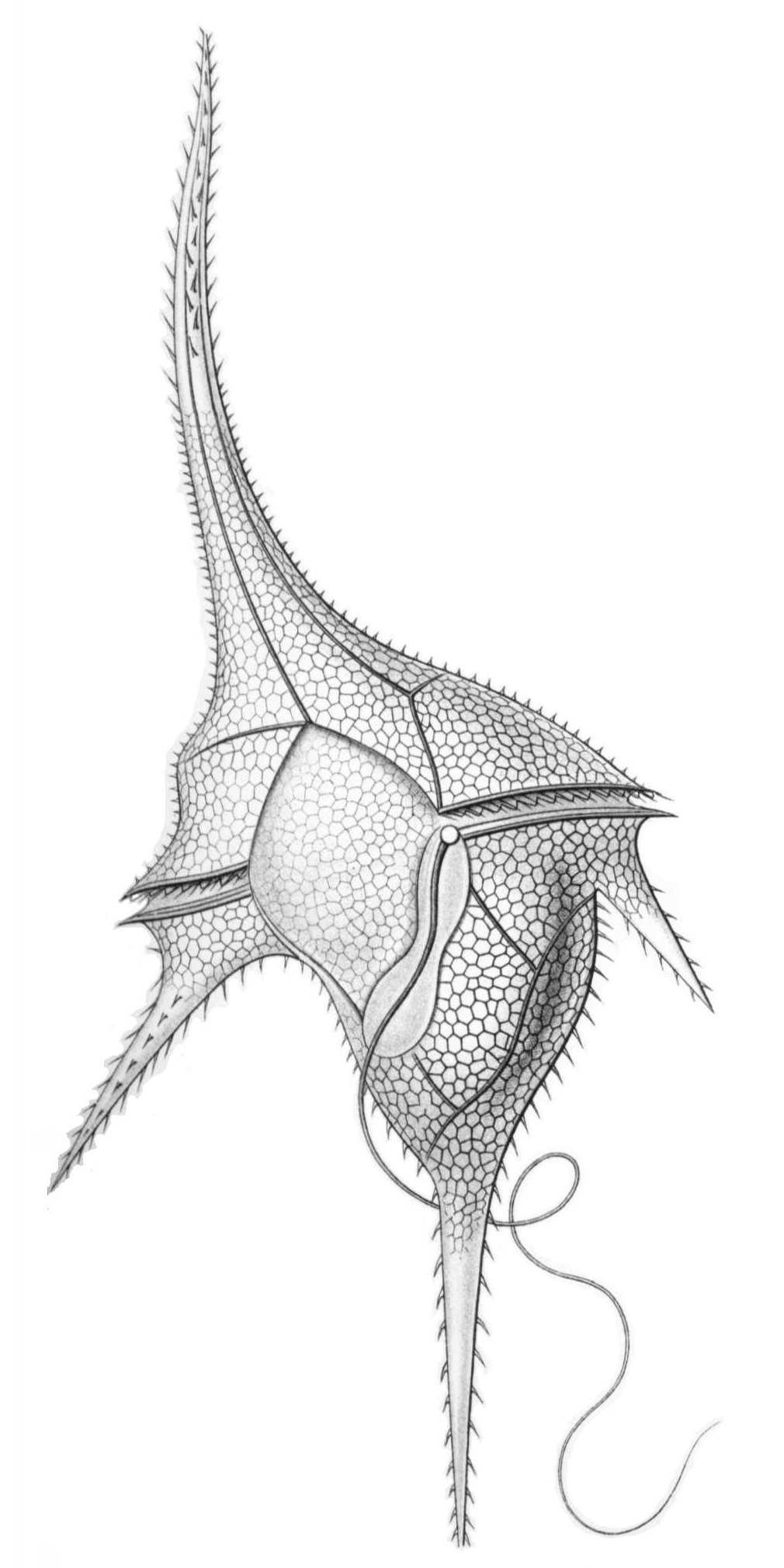
渦鞭毛藻類は一般に細胞に2本の鞭毛を持ち、一方は細胞の周囲に巻き付くように配置される横鞭毛(トランスバースフラジェラ)で、もう一方は前後に伸びる縦鞭毛(ロングチュディナルフラジェラ)です。この鞭毛の配置により、独特の回転運動(渦を巻くような運動)を行います。多くの種は細胞外側に厚い殻(テカ、セルロース板)を持つ「有殻型」と、殻を持たない「裸鞭毛藻」に分かれます。
光合成種はクロロフィルa・cやパリジニンなどの色素を持ち、海洋における一次生産に寄与します。一方で、捕食性や混合栄養性(光合成と捕食を併せ持つ)種も多く、栄養取得戦略は多様です。いくつかの種は硝酸塩やリンなどの無機栄養塩を効率的に利用し、低栄養環境で優位になります。
分類と系統学的特徴
従来の形態分類に加えて、分子系統学(SSU/LSU rDNA、ITSなど)により渦鞭毛藻類の分類は大きく見直されてきました。現在では、多くがアルベオラータ(Alveolata)という高次分類群に含まれ、同じグループには繊毛虫やアピコンプレックス(病原性原虫)が含まれます。系統や門・目の扱いは研究者によって異なり、プラスチド(葉緑体)の起源が複雑であることが分類の難しさを増しています。
葉緑体を持つ種の多くは、一次ではなく二次的または三次的な内共生(endosymbiosis)を経て様々な型の葉緑体を獲得しており、その起源は複数回にわたると考えられています。結果として、色素組成や葉緑体の膜構造が種によって大きく異なります。
生態(生態学的役割と行動)
渦鞭毛藻類は海洋・淡水の食物網で重要な役割を果たします。光合成種は一次生産者としてプランクトン群集の基盤を支え、捕食性種は微小動物や他の原生生物を捕食して物質循環に寄与します。また、多くの種が休眠胞子(シスト)を形成し、堆積物中で長期間生存して環境が良くなると再び出現することで個体群の維持に寄与します(古環境解析にも利用される「渦鞭毛藻シスト(dinocyst)」)。
一部の渦鞭毛藻は共生関係を形成します。代表例はサンゴの共生藻(かつてのゾキサンテラ、現在はSymbiodiniaceaeなど)で、ホストに光合成産物を供給することでサンゴ礁生態系を支えています。さらに生物発光を示す種(例:Noctiluca、Lingulodiniumなど)は夜間に光を放ち、捕食者の回避や餌の誘引などに関与する可能性があります。
赤潮・毒性と人間への影響
渦鞭毛藻の中には大量増殖(赤潮・有害藻類藻華)を起こす種があり、これは漁業・養殖業や沿岸生態系に大きな影響を及ぼします。いくつかの種は神経毒(サキシトキシンなど)やその他の毒素(ブレベトキシン、ドノトキシンなど)を産生し、貝毒(食中毒)や魚介類被害を引き起こします。赤潮は低酸素状態を招き海洋生物の大量死を招くこともあります。
分布要因と環境応答
渦鞭毛藻類の分布は、温度、塩分、光、栄養塩、海流および水深などの物理化学的要因に左右されます。河川流入や人為的な栄養塩負荷(富栄養化)が増えると、一部の有害種が優占しやすくなります。気候変動に伴う海水温の上昇や海洋の酸性化は種の分布や藻華の発生パターンを変える可能性があります。
研究手法と応用
渦鞭毛藻類は顕微鏡観察(光学・電子顕微鏡)、色素分析、分子遺伝学(DNAバーコーディング、メタバーコーディング)、培養実験などで研究されます。古環境復元や水質監視、赤潮予測モデルの構築、さらに共生藻研究はサンゴ礁保全や水産業への応用が期待されています。
まとめと今後の課題
渦鞭毛藻類は形態・生理・生態が非常に多様であり、海洋生態系で中心的な役割を持ちます。一方で分類学的な整理、葉緑体の起源解明、赤潮発生の予測と対策、生物多様性の把握(記載種と未記載種のギャップ解消)など、今後も多くの研究課題が残されています。
生物学
Zooxanthellae
渦鞭毛藻の中には褐虫藻と呼ばれる海洋真核生物の内共生体があり、その中にはサンゴなどの動物も含まれています。褐虫藻の中には、海中の真核生物の内共生体である褐虫藻があり、サンゴなどの動物が褐虫藻に依存している。褐虫藻を宿す生物としては、クラゲ、アサリ、有孔虫、ウミウシ、さらには繊毛虫や放散虫などがある。
レッドタイズ
渦鞭毛藻は、魚の毒となる「赤潮」の原因となります。渦鞭毛藻は、1mlあたり100万個以上の濃度で繁殖することもある。渦鞭毛藻の中には神経毒を産生するものもあり、このような量になると魚は死んでしまい、貝などの濾過性生物に蓄積され、それを食べた人間にも伝染する可能性があります。この現象は、ブルームが水に与える色から、赤潮と呼ばれている。しかし、すべての渦鞭毛藻が危険なわけではありません。夜の海で見られる青白い光は、生物発光性渦鞭毛藻のブルームによるものが多い。
分類
真核生物に分類されるとはいえ、渦鞭毛藻類の核は真核生物の核の主要な特徴を欠いている。渦鞭毛藻類の核は、原核生物のコイル状のDNA領域と、明確な真核生物の核との中間的な特徴を持っていることから、Dodgeはメソカリオンと呼んでいる。渦鞭毛藻の核は、原核細胞のコイル状のDNA領域と真核細胞の核の中間的な性質を持ち、メソカリオンと呼ばれている。
このグループには、多くの複雑なオルガネラや生活様式が含まれています。脊椎動物の目のような構造を持つもの、線虫を持つもの、プラスモディア(多核体)として生活するもの、2本の鞭毛を持つもの、光合成を行う渦鞭毛藻には様々な種類のプラスチドがあり、その遺伝学や細胞生物学は全体的にエキセントリックなものである。
化石
渦鞭毛藻類の化石は,地層の年代測定や相関関係を明らかにするための重要な資料である。渦鞭毛藻類はその一生の間に、移動可能なプランクトンの形態と、冬の間堆積物の中で生き延びることのできる抵抗性のあるシストの形態をとる。化石化されるのはシストの部分だけです。渦鞭毛藻類は、シルル紀に初めて出現し、中生代になってから大発生した。p440MargulisらはDinomastigotaという門に分類している。
関連ページ
質問と回答
Q: 渦鞭毛藻類とは何ですか?
A: 渦鞭毛藻類は、光合成を行うか、他の原生生物を捕食する従属栄養生物である鞭毛虫のグループです。
Q: 渦鞭毛藻類はすべて光合成をするのですか?
A: いいえ、光合成をするのは渦鞭毛虫の約半分だけで、残りは従属栄養性の捕食者です。
Q: 光合成をする渦鞭毛藻と捕食する渦鞭毛藻は、それぞれどのような名前で呼ばれているのですか?
A: 光合成を行う渦鞭毛藻類を「藻類」、捕食を行う渦鞭毛藻類を「原生生物」と呼ぶことがあります。
Q: 渦鞭毛藻類はどのような環境でよく見られるのですか?
A: 渦鞭毛藻類は、海洋プランクトンに多く生息していますが、淡水域にも多く生息しています。
Q: 渦鞭毛藻の個体数に影響を与える要因は何ですか?
A: 渦鞭毛藻の個体数は、海面水温、塩分濃度、水深の影響を受けます。
Q: 現在、自由生活する海洋渦鞭毛藻類は何種が報告されていますか?
A: 現在、約1,555種の海洋性自由生活渦鞭毛藻類が記載されています。
Q:渦鞭毛藻類は全部で何種類いると推定されていますか?
A: 最新の推定では、海洋性、淡水性、寄生性を合わせて2,294種の渦鞭毛藻類が生息しているとされています。
関連項目
著者
AlegsaOnline.com 渦鞭毛藻類とは|特徴・分類・生態 — 光合成・捕食・分布の解説 Leandro Alegsa
URL: https://ja.alegsaonline.com/art/27497
出典
- cicimar.ipn.mx : "A checklist and classification of living dinoflagellates (Dinoflagellata, Alveolata)"
- tolweb.org : Tree of Life project