種分化(スペシエーション)とは:定義・原因・アロパトリと交配の役割
種分化の定義・原因を解説。アロパトリと交配が新種形成に果たす役割や最新研究の遺伝子流動をわかりやすく紹介
種分化とは、種が新たに形成される過程のことです。これは進化生物学の中心的な問題の一つであり、どのようにして多様な生物種が生まれるのかを理解するための基本概念です。
歴史的な見解と用語
ダーウィンは、多くの種が既存の種から連続的に変化して生じると考えました。彼の考え方では、ある系統が時間をかけて変化し続けることで新しい形質を獲得し、最終的に元の集団と十分に異なれば別の種と見なされます。この過程は一般にアナゲネシス(anagenesis)、あるいは「系統内進化」と呼ばれます。
20世紀には、別のプロセスがより重要視されるようになりました。多くの生物学者は、既存の種が分裂して二つ以上の独立した系統になる現象、すなわちクラドゲネシス(分岐的種分化)が種の起源を説明すると考えました。ここでは、集団間の隔離や分離メカニズムが鍵となるという見方が支配的でした。
画像ギャラリー
10 画像









物理的分離(アロパトリ)とそのメカニズム
長年、地理的隔離(アロパトリック種分化)が新種の発生における主因であると考えられてきました。これは元々一緒に暮らしていた集団が地理的に分断され、遺伝子流動(個体の移動や交雑)がおこなわれなくなることで、各集団で別々の進化が進行するというモデルです。
物理的分離を引き起こす代表的なプロセスには以下があります:
- 分断(vicariance): 地殻変動、気候変化、海面上昇などにより利用可能な生息域が分割される。
- 分散(dispersal): 一部の個体が隔たった新しい領域に移住し、小さな創設集団が新たな進化を始める(末端隔離/peripatric)。
地理的隔離下では、遺伝的漂流や自然選択、性的選択がそれぞれの集団で異なる方向に働き、異なる形質や生殖的隔離を生み出します。やがて、交配しても生殖的に不成功(あるいは不利)となるほどの違いが蓄積されれば、別種として認められます。
生殖的隔離とその種類
種分化の核心はしばしば生殖的隔離です。これには主に以下の二種類があります:
- 前接合的隔離(prezygotic): 時間的隔離(繁殖期のずれ)、生態的隔離(異なる生息地)、行動的隔離(求愛行動の違い)、機械的隔離(交尾器の不適合)、配偶子レベルの隔離(受精の失敗)など。
- 後接合的隔離(postzygotic): 雑種の不妊や低適応度(雑種不適合)、雑種の発生異常など。
これらの隔離は単独で起こることも、複数が組み合わさって種分化を完了させることもあります。隔離が不完全な段階では交雑帯(hybrid zone)が形成されることがあり、そこでは遺伝子のやり取りが続きます。
交配(ハイブリディゼーション)とゲノム解析が示す新しい見方
過去20年ほどの分子生物学・ゲノム解析の進展により、近縁種間でしばしば交配がおこり、その結果として遺伝子が一方向または双方向に移動(遺伝子浸透(introgression))することが明らかになってきました。これは従来の「完全に隔離された種」モデルを修正する必要を示しています。
重要な点:
- 交配があるからといって必ずしも種の区別が無意味になるわけではない。多くの場合、種は一部の遺伝領域では混ざり合う一方、他の重要な遺伝領域では隔離が保たれる「ゲノムの島(genomic islands of divergence)」のようなパターンを示します。
- 交配を通じて有利な遺伝子が種間で伝わることがあり、これが適応に寄与する場合がある(例:病原抵抗性や環境耐性の獲得)。
- 一部の雑種が新種として成立することもある(雑種新種形成(hybrid speciation))。特に植物では倍数化(ポリプロイディ)を伴うケースが多く、これにより速やかに生殖隔離が成立します。
アロパトリ以外の種分化様式
地理的隔離以外にも種分化が起こることが知られています:
- 同所的種分化(sympatric speciation): 同一地域内で生息域や資源利用の差、強い選択圧や生殖的選好の変化により新種が生じる。例として寄生昆虫や食性差による分化などが議論されてきました。
- 傍接的種分化(parapatric speciation): 隣接する生息地間で連続的な環境差があり、境界付近で適応と部分的隔離が進むことで種分化が進行する場合。
まとめ — 現代の理解
現代の種分化の理解は、単一の単純なモデルに還元されるものではなく、複数の力(地理的隔離、自然選択、遺伝的漂流、性的選択、交配・遺伝子浸透など)が相互に作用する複合的な過程と捉えられています。重要なのは、生殖的隔離がどのようにして成立するかと、それがゲノムレベルでどのように現れるかを理解することです。
最後に、種分化の研究は観察、実験、ゲノム解析を組み合わせることで進展しており、これにより「種」という概念そのものの定義や適用の仕方についても柔軟で現実的な見方が広がっています。
隔離のメカニズム
隔離機構とは、種の中でグループ間の繁殖が成功しないようにするものです。集団の生殖的隔離が確立される。これは生物学的な種の概念にとって特に重要であり、種は生殖的隔離によって定義される。
隔離機構は、受精前と受精後の2つのグループに分けられます。
受精前
個体の交配を妨げる要因。
- 地理的隔離。異なる地域に生息する種で、しばしば障壁で隔てられている。
- Temporal isolation。個体の活動時間が異なるため、交尾をしない。これは1日のうちの異なる時間帯であったり、異なる季節であったりします。種の交尾期間が一致しない場合もある。交尾期間中、あるいは全く交尾しない個体もある。
- 生態学的隔離。個体は自分の好む生息地でのみ交尾する。生態的に異なる嗜好を持つ他の種の個体と出会うことはない。
- 行動的隔離。異なる種の個体が出会うことはあっても、性的な合図があってもそれを認識しない。ほとんどの場合、個体は自分の種のメンバーを選ぶ。
- 機械的に隔離する。交尾を試みても、精子の移動が行われないことがある。個体の大きさや形態が原因で相容れない場合がある。
- 胎児の不適合。精子の授受は行われるが、卵子は受精しない。
受精後
遺伝子の不適合、ハイブリッドの不成立、不稔など、交配が成功しない要因。
- 接合体死亡率。卵は受精したが、接合体が発生しないこと。
- Hybrid inviability:ハイブリッド胚は形成されるが、生存能力はない。
- 雑種不稔:雑種は生存可能だが、生まれた成虫は不稔である。
- ハイブリッドの故障。第一世代(F1)の雑種は生存しており、肥沃であるが、それ以上の世代(F2や戻し交配)の雑種は生存していないか不稔である。

地理的な孤立
これが種分化の最も一般的な原因だと考えられている。これを最初に考えたのは、ドイツの探検家・博物学者であるモーリッツ・ワグナーである。
ワグナーの初期のキャリアは地理学者としてのもので、北アフリカ、中東、熱帯アメリカについての地理書を数多く出版した。彼はまた、熱心な博物学者、収集家でもあり、生物学者の間ではこの仕事で最もよく知られている。進化論者であり、生物学の歴史家でもあるエルンスト・マイヤーは、ワグナーの意義について次のように述べている。 p562–565
アルジェリアに滞在した3年間、ワグナーは飛べない甲虫のPimeliaとMelasomaを研究しました。それぞれの属にはいくつかの種があり、それぞれの種はアトラス山脈から地中海へと下る川の間にある北海岸の一帯に限られている。川を渡るとすぐに、異なるが近縁の種が現れる。
「初期の種は、少数の個体がその範囲の限界を超えたときにのみ(発生する)......新しい種の形成は、入植者が他の種のメンバーから長期間継続して分離しない限り......決して成功しない」。
これは、ある種の地理的種分化のプロセスを初期に記述したものでした。1942年にマイヤーによって再び紹介され、地理的種分化の重要性が進化論的統合の中核的な考えの一つとなった。
地理的種分化の別の用語として、アロパトリス・スペシエーション(allopatric speciation)があります。allopatryとは「異なる土地」という意味です。
空き島
火山島は生物がいない状態で形成されており、すべての生物は風や水に運ばれてやってくるしかありません。ハワイ諸島やガラパゴス諸島では、本土から島にたどり着いた時点で、すべての生命体が変化することがわかっています。
約6,500平方マイル(約17,000km2)のハワイ諸島には、熱帯雨林から山の牧草地まで、世界で最も多様なショウジョウバエが生息していると言われています。ハワイのショウジョウバエは約800種が知られている。
研究によると、古い島から新しい島への種の明確な「流れ」が見られます。また、古い島に戻って植民地化したり、島を飛び越えたりするケースもありますが、頻度ははるかに低いです。
カリウム/アルゴン放射性物質による年代測定では、現在の島々は0.4百万年前(マウナケア)から10百万年前(ネッカー)にさかのぼる。ハワイ諸島の中で、現在も海の上にある最古の島はクレ環礁で、その年代は30百万年前とされています。
太平洋プレートがホットスポットの上を移動することで生まれたこの列島自体は、はるか昔、少なくとも白亜紀には存在していたのだ。ハワイ諸島と海中にある旧島々は、ハワイ・エンペラー海山列島を構成しており、海中の山の多くはガイオットである。
ハワイに生息するショウジョウバエのすべての種は、約2000万年前に島に入植した単一の祖先種の子孫であることが明らかになっている。その後の適応放散は、競争相手がいないことと、多様なニッチが空いていることによって促進された。一人の妊娠した女性が島を植民地化した可能性もありますが、同じ種のグループであった可能性の方が高いと思われます。
ハワイ諸島には他にも同じような適応放散をした動物や植物がありますが、その数はそれほど多くありません。
リング種
生物学では、環状種とは、隣り合う個体群がつながった一連のものであり、それぞれの個体群が隣り合う個体群と交配することができる。鎖の両端が重なっている。
このシリーズの2つの末端集団は、交配するにはあまりにも遠い関係にある。このように、遺伝的にはつながっているものの、交配していない「末端」集団が同じ地域に共存することで、「環」を閉じている場合があります。
環状種は進化の重要な証拠であり、集団が遺伝的に分岐することで時間的に何が起こるかを示している。リチャード・ドーキンスは、環状種は「時間の次元では必ず起こることを、空間の次元でのみ示している」と述べている。
しかし、シンプルでわかりやすい例はなかなかありません。
カスミチョウチョウウオ
環状種の典型的な例は、Larus gullsのcircumpolar species ringである。これらのカモメの範囲は、北極点を中心としたリングを形成しているが、通常、個々のカモメが飛び交うことはない。
大まかに言えば、リングの両端を除いて、カモメは隣人とある程度交配することができます。
カモメのグループは、ヨーロッパで2つの系統が出会う場所を除き、連続体を形成しています。
現実はもっと複雑ですが、一般的な考え方をよく表しています。
エンサティナ・サンショウウオ
エンサティナサンショウウオは、カリフォルニアのセントラルバレー周辺の山に生息する環状種。山の周りには馬蹄形の複合体が形成されています。馬蹄の周りにある19の各個体群の間では交配が起こりうるが、馬蹄の西端にあるEnsatina eschscholtzii亜種は、東端にあるEnsatina klauberiと交配することはできない。種分化の過程のほぼすべての段階」(ドブジャンスキー)を示しています。リチャード・ハイトンは、エンサティナは複数の種が存在するケースであり、1つの種が連続しているわけではないと主張した。
ミドリムシムシムシムシムシムシ
ミドリムシクイ(Phylloscopus trochiloides)にはいくつかの亜種がありますが、その中でもヨーロッパで最もよく知られているのがP. t . viridianusです。この種は、チベット高原の東西で個体群が分岐し、後に北側で合流する環状種である。その関係はかなり混乱している。




共時的な種分化
同所的種分化とは、1つの祖先種から2つ以上の子孫種が形成され、それらがすべて同じ地理的場所に存在すること。現在では非常によく見られる現象だと考えられている。
同所的種分化とは、同じ場所に生息しながら種が分岐すること。同所的種分化の例としてよく挙げられるのは、同じ地域で異なる宿主植物に依存するようになった昆虫である。
種分化のメカニズムとしての同所的種分化の存在については、熱い議論が交わされた。同所的種分化の証拠は、実際にはミクロ地理的種分化の例であると主張する人々がいました。一般的には、これは現在では好ましい説明ではありません。むしろ、近縁種同士の交配の結果、そのような交配の子孫に自然淘汰が働いたのだと考えられている。同所的種分化の例としては、東アフリカのナブガボ湖のシクリッドが広く知られているが、これは性淘汰によるものと考えられている。
倍数体化による種分化
例えば、4倍体×2倍体の交配では、しばしば3倍体の不稔子孫が生まれるため、多倍体化は多くの急速な種分化を引き起こしてきた。
しかし、すべての倍数体が親植物から生殖的に隔離されているわけではなく、例えば、3倍体のハイブリッドと2倍体の交配で4倍体が生まれるなど、遺伝子の流れが発生することもあります。
現存する植物種や動物種の多くは、その進化の過程で明らかに倍数体化を経験しています。成功した倍数体種の生殖は、時には無性生殖、つまり単為生殖によって行われます。理由は不明だが、多くの無性生物は倍数体である。
サンザシフライ
進化の一例として、サンザシバエ(Rhagoletis pomonella)の場合、同所的種分化が進んでいるように見えます。
サンザシバエの個体群は、それぞれ異なる果実を食べます。19世紀の北米では、外来種であるリンゴが導入されてからしばらくして、異なる個体群が出現しました。このリンゴを食べる集団は、通常はリンゴだけを食べ、歴史的に好まれてきたサンザシの果実は食べない。現在のサンザシを食べる集団は、通常はリンゴを食べません。
同所的な種分化が起こっていることを示唆する証拠もある。サンザシのハエはリンゴのハエに比べて成熟する時期が遅く、成熟するまでに時間がかかります。また、交配の証拠はほとんどありません(研究者は交配率が4〜6%であると記録しています)。
新しいサンザシのフライの出現は、進化が進んでいることを示す例です。
ハイブリッド化
稀に、異なる種の個体が交配して新種が生まれることがある。通常、そのような交配の産物は不妊であるか、あるいは比較的不妊であるため、自然選択によって淘汰されてしまう。
新しいハイブリッド種の成功例が一つ見つかった。この新種は、イタリアスズメとスペインスズメの交配種で、血液中のDNAからの塩基配列分析により、基本的な事実が確認された。この鳥は、イタリアの両方の親種が住んでいる地域に生息している。並んで暮らしていても、スペインのスズメとは繁殖しない。
人工的な種分化
家畜化された動物の飼育によって新種が作られてきたが、最初の時期や方法ははっきりしていない。例えば、家畜のヒツジは雑種によって作られたものであり、ヒツジの子孫であるOvis orientalisとの間にはもはや子孫を残すことができない。
一方、家畜の牛は、野生の牛、ガウア、ヤクなどの数種類の品種と、容易に繁殖可能な子孫を作ることができるので、同じ種と考えてよいでしょう。
ラボ種
実験室での新種誕生は、1980年代後半に行われたものが最もよく知られている。ウィリアム・ライスとG.W.ソルトは、明暗や湿潤・乾燥といった3種類の生息環境を選択できる迷路を用いて、ショウジョウバエを飼育しました。
各世代を迷路の中に入れ、8つの出口のうち2つの出口から出てきたハエのグループは、それぞれのグループ内でお互いに繁殖するように分けた。35世代後、2つのグループとその子孫は、生息地の選好性が強いため、生殖的に隔離された。彼らは自分たちが好むエリア内でのみ交尾し、他のエリアを好むハエとは交尾しなかったのである。このような試みの歴史は、Rice and Hostert (1993)に記載されています。
また、ダイアン・ドッドは、ショウジョウバエの交尾の好みから、生殖隔離がどのように発展するかを、デンプンとマルトースという異なる種類の食物を用いて、わずか8世代後に示すことができました。
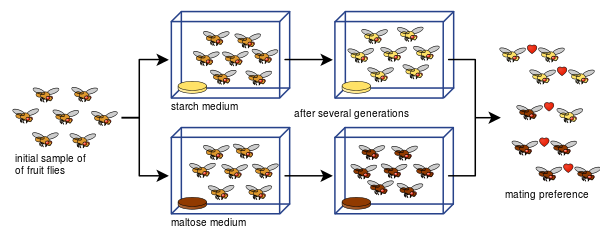
ドッドの実験は、他の種類のミバエや食品でも、多くの人が簡単に再現することができる。
強化(ウォレス効果)
強化とは、自然淘汰が生殖的分離を高めるプロセスである。
同じ種の2つの個体群が分離した後、再び接触したときに起こることがあります。もし、繁殖上の隔離が完全であれば、すでに2つの相容れない別の種に発展しているはずです。
生殖的隔離が不完全であれば、集団間の交配が進めば雑種が生まれ、その雑種が不妊である場合もある。その雑種が不妊であったり、不妊であっても祖先よりも体格が劣っていたりする場合は、それ以上の生殖的隔離はなく、基本的に種分化が起こっていることになります(例:馬やロバのように)。
その理由は、もしハイブリッドの子供の親がそれぞれ異なる環境で自然に選択された形質を持っているならば、ハイブリッドの子供は両方の形質を持ち、どちらの親よりもどちらのニッチにも適合しないだろうということである。雑種の体力が低下すると、選択は同種交配を好むようになり、雑種化が減少することになる。
これは、19世紀末に進化生物学者のアルフレッド・ラッセル・ウォレスが種分化の重要な要因ではないかと指摘したことから、ウォレス効果と呼ばれることもある。
もしハイブリッドの子孫が祖先よりもフィットしていれば、その個体群は接触している地域内で再び同じ種に融合する。
ある種の2つの形態の間に「ハイブリッドゾーン」がある場合、分離のための補強が必要です。ハイブリッドゾーンとは、分岐した個体群が出会い、交配する地域のことである。このような地域ではハイブリッドの子孫が非常によく見られ、これは通常、分岐した種が二次的に接触することで生まれる。
強化がなければ、2つの集団や種は制御不能な交配をしてしまいます。強化は、上述のように人工的な選択実験で誘発することができる。
質問と回答
Q:種分化とは何ですか?
A:種分化とは、種がどのように形成されるかという過程のことです。進化生物学の主要な部分である。
Q:ダーウィンは種の形成についてどう考えていたのか?
A:ダーウィンは、ほとんどの種は既存の種から直接生まれたと考えており、これを「無原罪化」あるいは「植物的進化」と呼んでいます。
Q: 20世紀には、種分化は一般的にどのように捉えられていたのでしょうか?
A: 20世紀には、ほとんどの種は、先に存在した種が分裂してできたものだと考えられていた。これは「クラドジェネシス」と呼ばれていた。また、この分裂は、物理的な分離などの分離メカニズムによって引き起こされたり、助けられたりすると考えられていた。
Q: 最近の研究によって、種分化についての理解はどのように変わったのでしょうか?
A: 最近の研究では、近縁種同士の交配が起こりうること、そしてその交配によって遺伝子が伝達されうることが明らかになっています。つまり、種の定義に生殖的な隔離は必ずしも必要ではなく、種分化に同系交配(物理的な分離)は必要ない可能性があるのです。
Q:物理的な分離と新種の形成はどう関係するのですか?
A: かつて共存していた種が物理的に分離することが、新種形成の主な要因であると考えられており、科学文献で取り上げられている多くの例で説明されています。
関連項目
著者
AlegsaOnline.com 種分化(スペシエーション)とは:定義・原因・アロパトリと交配の役割 Leandro Alegsa
URL: https://ja.alegsaonline.com/art/92570
出典
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833